
Sill/Strömming
Kartan visar de områden där arten förekommer. Observera att detta kan skilja sig från de områden där bedömningar har gjorts för arten.
Karta över hav och sjöar
Karta över Ices-områden
Sill/Strömming 2022
Bestånds- och populationsstruktur
Internationella havsforskningsrådet (Ices) har identifierat sex olika bestånd av sill/strömming i Östersjön och Nordsjön. Beslutet är en kompromiss mellan att separat behandla alla de sillpopulationer som har beskrivits på biologiska grunder under tid, och de praktiska begränsningar som finns i form av områden för fångstrapportering och möjlighet att korrekt hänföra enskilda fiskar till en viss population. I Bottniska viken betraktas Bottenviken och Bottenhavet som ett bestånd sedan 2017. Två bestånd behandlas i centrala Östersjön, ett i Ices-delområden 25–29 och 32 samt ett i Rigabukten (Ices-delområde 28.1 som inte är inkluderat i denna rapport). Sillen i sydvästra Östersjön (Ices-delområden 22–24) behandlas tillsammans med vårlekande sill i Kattegatt och Skagerrak på grund av sitt vandringsbeteende. Sillen har tidigare förvaltats som två enheter med två separata kvoter, en för hela Egentliga Östersjön (Ices-delområden 22–28, 29 södra och 32) och en för Ices-delområden 29 norra, 30 och 31. År 2005 ändrades förvaltningsenheterna så att de överensstämmer med Ices beståndsindelning. Generellt är sillen i norra Östersjön mer långsamväxande och har lägre medelvikt per ålder än sillen i södra Östersjön. Medelvikten har minskat det senaste decenniet på grund av olika miljöfaktorer och eventuellt som resultat av konkurrens inom arten¹. Därutöver finns ytterligare två sillbestånd: höstlekande sill i Nordsjön, Skagerrak, Kattegatt och östra Engelska kanalen samt det norska vårlekande sillbeståndet i Nordsjön, Skagerrak och Kattegatt. Beståndsuppdelningen av sill och strömming i Östersjön har framför allt i Sverige under senaste året blivit ifrågasatt, i och med rapporter från fisket om en minskning av stor strömming vid den svenska kusten. Forskningen visar att sillen och strömmingen generellt blivit mindre under senare tid och når könsmognad vid tidigare ålder (WGBFAS 2022), men hur minskningen av stor strömming på kusten kan kopplas till fiskets mönster och beståndens uppdelningar i separata eller blandade lekkomponenter är oklart. I en ny studie av Fan Han et al. (2020)³² har man genom att sekvensera hela genomet av sill och strömming visat genetiska skillnader mellan olika lekkomponenter, som grundar sig i anpassningar av individer till lokala miljöförhållanden såsom temperatur, salinitet och ljusförhållanden. Tekniken att identifiera så fint upplösta genetiska skillnader öppnar för en möjlighet att kartlägga beståndsuppdelningen av strömming och sill på ett sätt som inte varit möjligt tidigare. En insamling och genetisk analys av individer från olika lekansamlingar pågår längst med den svenska östkusten (Stockholms Universitet och Uppsala Universitet i samarbete med SLU Aqua) som en start för att urskilja gränsdragningen mellan bestånd i kustområdena.
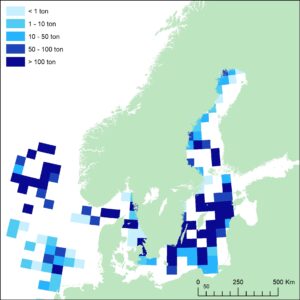
Figur 1. Svenska yrkesfiskares huvudsakliga landningar (ton) av sill/strömming 2021 per Ices-rektangel. En Ices rektangel är cirka 56 km x 56 km stor.
Biologiskt råd för sill/strömming i centrala Östersjön utom Rigabukten
Internationella havsforskningsrådet (Ices)
Ices fångstråd för sill/strömming i centrala Östersjön (Ices-delområden 25–29 och 32) utom Rigabukten för 2023 är mellan 70 130 och 95 643 ton³. För 2022 var rådet mellan 52 443 och 87 581 ton. Jämfört med 2022 innebär rådet en ökning med 33 procent av de rekommenderade fångstmängderna.
Institutionen för akvatiska resurser (SLU Aqua)
SLU Aquas råd för 2023 följer Ices rådgivning.
ICES råd om fiskemöjligheter i Centrala Östersjön (Ices-delområden 25-27, 28.2, 29 och 32).
Biologiskt råd för strömming i Bottniska viken
Internationella havsforskningsrådet (Ices)
Ices fångstråd för strömming i Bottniska viken (Ices-delområden 30–31) för 2023 är mellan 80 047 och 103 059 ton14. För 2022 var rådet mellan 86 729 och 111 714 ton. Jämfört med 2022 innebär rådet en minskning med 7,7 procent av de rekommenderade fångstmängderna.
Institutionen för akvatiska resurser (SLU Aqua)
SLU Aquas råd för 2023 följer Ices rådgivning.
ICES råd om fiskemöjligheter i Bottniska viken.
Biologiskt råd för vårlekande sill i Skagerrak, Kattegatt och sydvästra Östersjön
Internationella havsforskningsrådet (Ices)
Ices fångstråd för vårlekande sill i Skagerrak, Kattegatt och sydvästra Östersjön (Ices-delområden 20–24) samt Nordsjön (Ices-område 4) för 2023 är noll ton19. För 2022 var rådet noll ton. Jämfört med 2022 är rådet oförändrat.
Institutionen för akvatiska resurser (SLU Aqua)
SLU Aquas råd för 2023 följer Ices rådgivning.
ICES råd om fiskemöjligheter i Skagerrak, Kattegatt och sydvästra Östersjön samt Nordsjön (Ices-delområden 20–24 samt Ices-område 4).
Biologiskt råd för höstlekande sill i östra Engelska kanalen, Nordsjön, Skagerrak och Kattegatt
Internationella havsforskningsrådet (Ices)
Ices fångstråd för höstlekande sill i östra Engelska kanalen, Nordsjön, Skagerrak och Kattegatt för 2023 är 414 886 ton22. För 2022 var rådet 532 183. Jämfört med 2022 innebär rådet en minskning med 22 procent av de rekommenderade fångstmängderna.
Institutionen för akvatiska resurser (SLU Aqua)
SLU Aquas råd för 2023 följer Ices rådgivning.
ICES råd om fiskemöjligheter i Nordsjön, Skagerrak, Kattegatt och östra Engelska kanalen.
Biologiskt råd för norsk vårlekande sill i nordöstra Atlanten och Arktis
Internationella havsforskningsrådet (Ices)
Ices fångstråd för norsk vårlekande sill för 2023 är 511 171 ton31. För 2022 var rådet 598 588 ton. Jämfört med 2022 innebär rådet en minskning med 15 procent av de rekommenderade fångstmängderna.
Beskrivningen av norsk vårlekande sill är inte kopplad till ett specifikt område och Ices råd gäller för alla områden där sillen från detta bestånd befinner sig. Rådet är i enlighet med den av EU, Färöarna, Island, Norge och Ryssland överenskomna förvaltningsplanen.
Institutionen för akvatiska resurser (SLU Aqua)
SLU Aquas råd för 2023 följer Ices rådgivning.
ICES råd om fiskemöjligheter i Nordöstra Atlanten och Arktis.
Sill/Strömming 2022
Havsfiskelaboratoriet
Sida publicerad: 8 april 2022